产品









服务
联系我们
400-021-7882
021-54281913
021-54281913
|
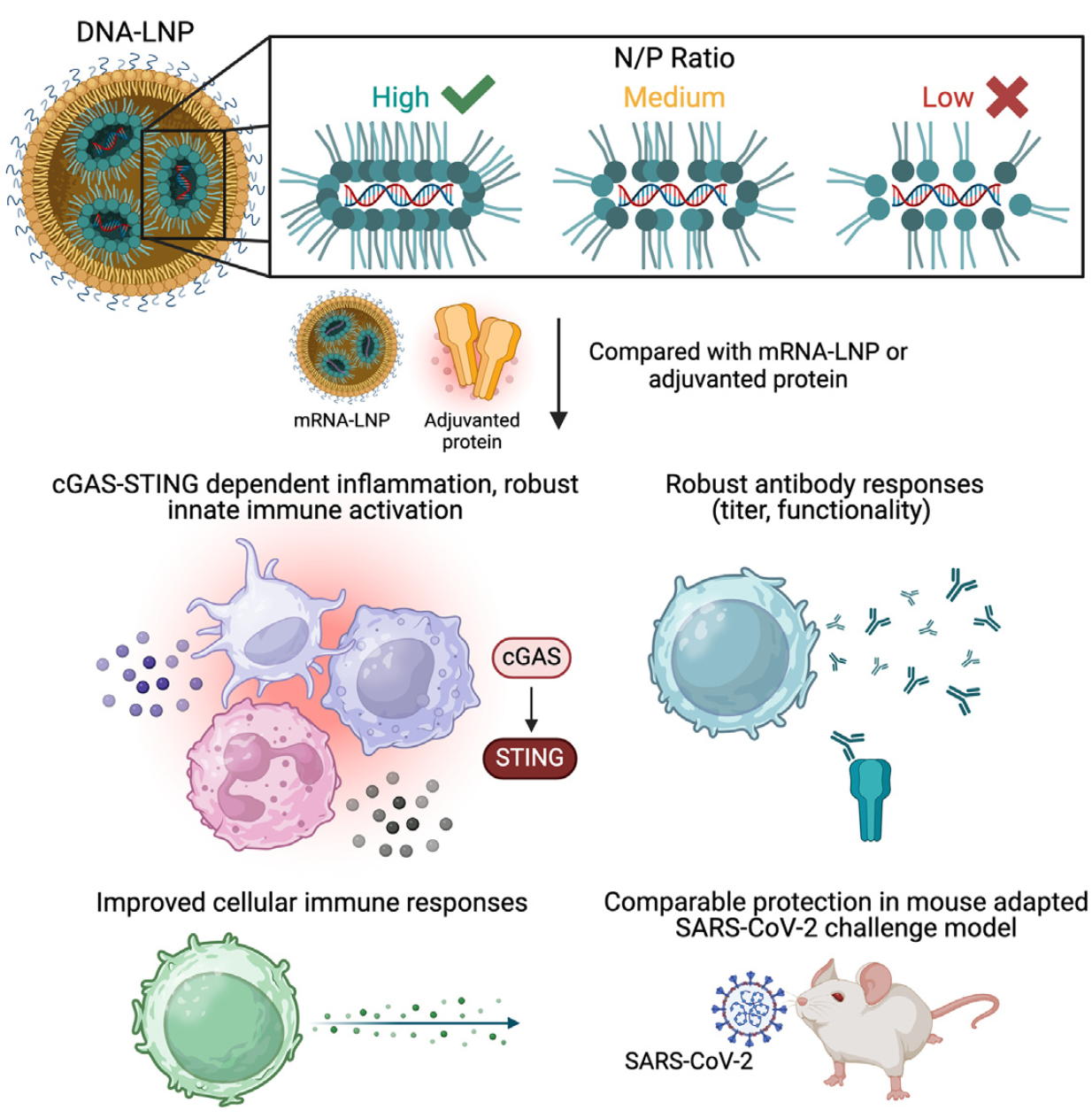
疫苗是抵御病原体感染的有力武器,能够激发适应性免疫的疫苗尤为重要。核酸疫苗,包括mRNA疫苗和DNA疫苗,因其独特优势在近年来备受关注。mRNA疫苗在COVID-19预防中表现出色,如Moderna的SpikeVax和Pfizer-BioNTech的Comirnaty已获批使用。然而,mRNA疫苗在生产和储存方面存在挑战,如需要冷链运输。DNA疫苗则具有临床安全性高、稳定性好且生产简单的优点,但裸DNA免疫原性较差。近期,美国费城wistar研究所疫苗和免疫治疗中心David B. Weiner团队在Cell Reports Medicine发表名为“Modulation of lipid nanoparticle-formulated plasmid DNA drives innate immune activation promoting adaptive immunity”的研究。本研究旨在探索脂质纳米颗粒包裹的质粒DNA(DNA-LNP)疫苗,以期发挥其优势,克服现有问题。 |
|||||
|
示意图 |
|||||
结果
|
|||||
| 1. 脂质成分与DNA比例对免疫原性的影响: | |||||
|
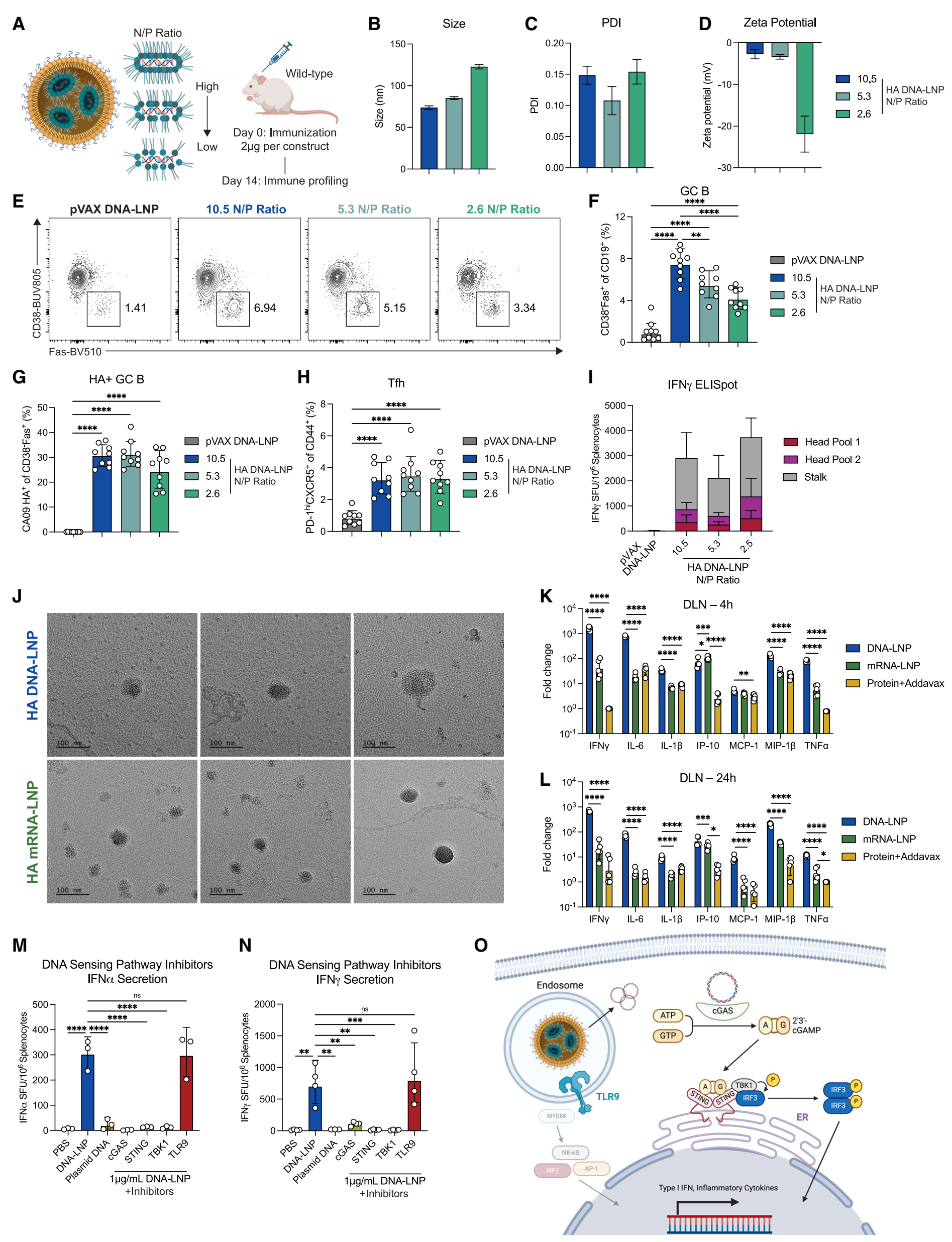
研究制备了不同N/P比例(10.5、5.3、2.6)的CA09血凝素(HA)表达质粒DNA-LNP(HA DNA-LNP),并考察了其生物物理特性和免疫原性。表征显示,10.5和5.3 N/P比例的DNA-LNP具有相似的封装效率(分别为91%和89%),而2.6 N/P比例的封装效率略低,接近80%。2.6 N/P比例的DNA-LNP颗粒较大,zeta电位更低。免疫小鼠后,发现10.5和5.3 N/P比例的DNA-LNP能显著提高总生发中心(GC)B细胞的频率,而2.6 N/P比例则显著降低。尽管如此,所有N/P比例的DNA-LNP均能诱导相似的激活T滤泡辅助细胞(Tfh)反应和抗原特异性CD4+及CD8+ T细胞反应。 透射电子显微镜(TEM)结果显示DNA-LNP和mRNA-LNP在形态上相似。体内成像显示,Luciferase DNA-LNP在注射后40天内持续表达于肌肉中,而Luciferase mRNA-LNP信号在表达后迅速减弱至基线水平,且在肝脏中短暂表达。 |
|||||
| 2. DNA-LNP通过cGAS-STING通路诱导炎症反应: | |||||
|
局部促炎细胞因子产生:免疫后4小时和24小时,与PBS对照组相比,HA DNA-LNP和HA mRNA-LNP均显著上调了多种促炎细胞因子,如IFNγ和IL-6。mRNA-LNP在4小时时诱导了较高的IP-10上调,而DNA-LNP在24小时时IP-10水平与之相当。 DNA感知通路的机制研究:使用化学抑制剂阻断cGAS-STING-TBK1通路后,DNA-LNP诱导的IFNα和IFNγ分泌显著减少,而阻断TLR9通路则无此效果。这表明cGAS-STING通路是DNA-LNP诱导炎症反应的主要驱动因素。 |
|||||
|
Figure 1. Initial immune characterization of HA DNA-LNP formulations and immunogenicity |
|||||
| 3. HA DNA-LNP诱导强烈的早期适应性和先天免疫激活: | |||||
|
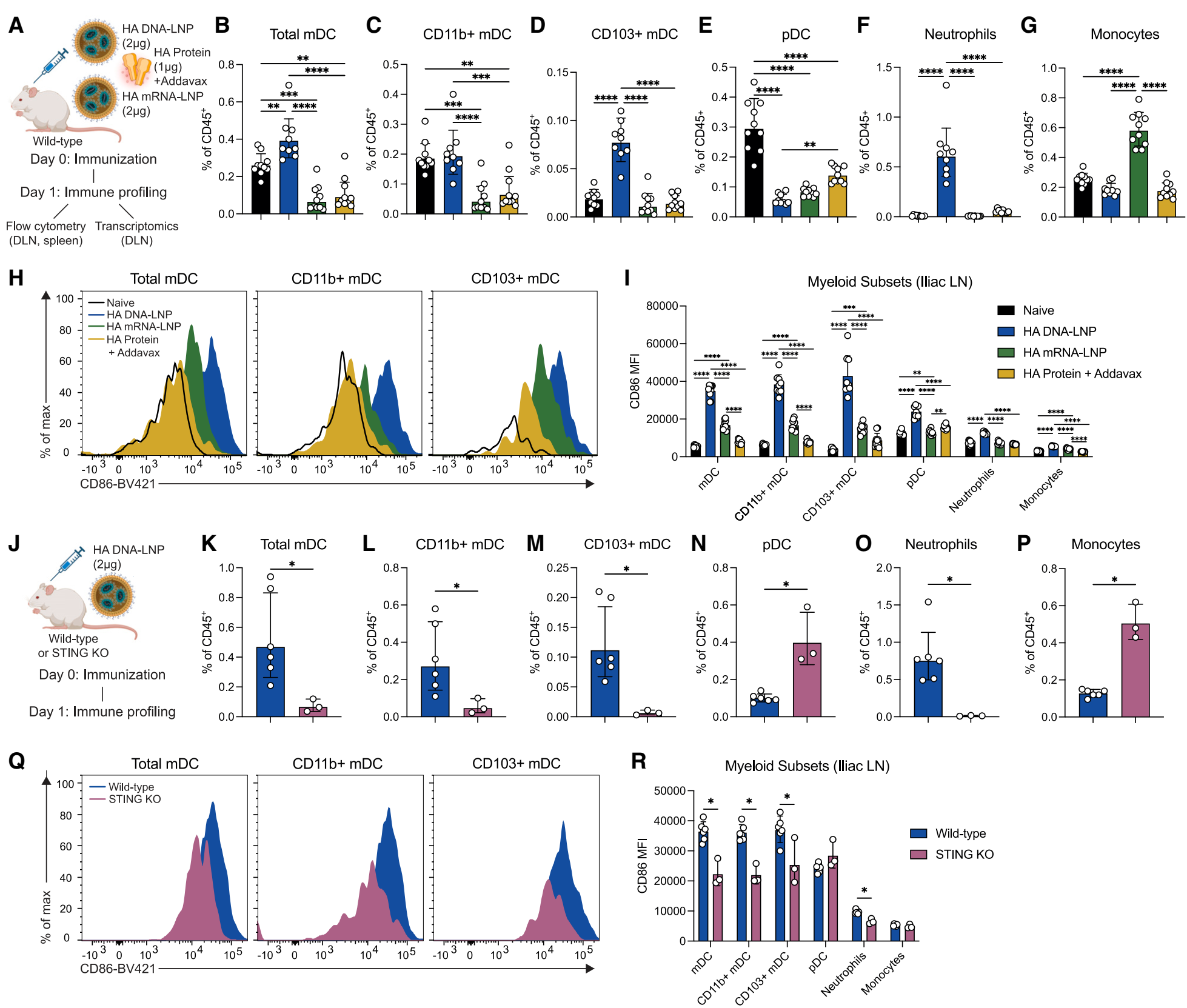
免疫细胞亚群的变化:免疫后24小时,与mRNA-LNP和佐剂蛋白相比,DNA-LNP显著增加了迁移性树突状细胞(mDCs)在引流淋巴结(DLNs)中的频率,特别是CD103+ mDCs。同时,DNA-LNP还显著增加了中性粒细胞的频率,而mRNA-LNP则增加了单核细胞的频率。 免疫细胞激活状态:DNA-LNP免疫诱导了所有髓系细胞亚群的激活,特别是mDCs和CD103+ mDCs的CD86表达显著增加。在脾脏中,DNA-LNP免疫也诱导了类似的髓系细胞激活模式,但mDCs频率保持相对稳定。 cGAS-STING依赖性验证:在STING敲除小鼠中,DNA-LNP诱导的髓系细胞频率和激活状态显著降低,表明这些反应依赖于cGAS-STING通路。 |
|||||
|
Figure 2. Early immune profiling of innate and adaptive populations reveals STING-dependent activation status after HA DNA-LNP immunization |
|||||
| 4. HA DNA-LNP诱导强烈的T细胞反应: | |||||
|
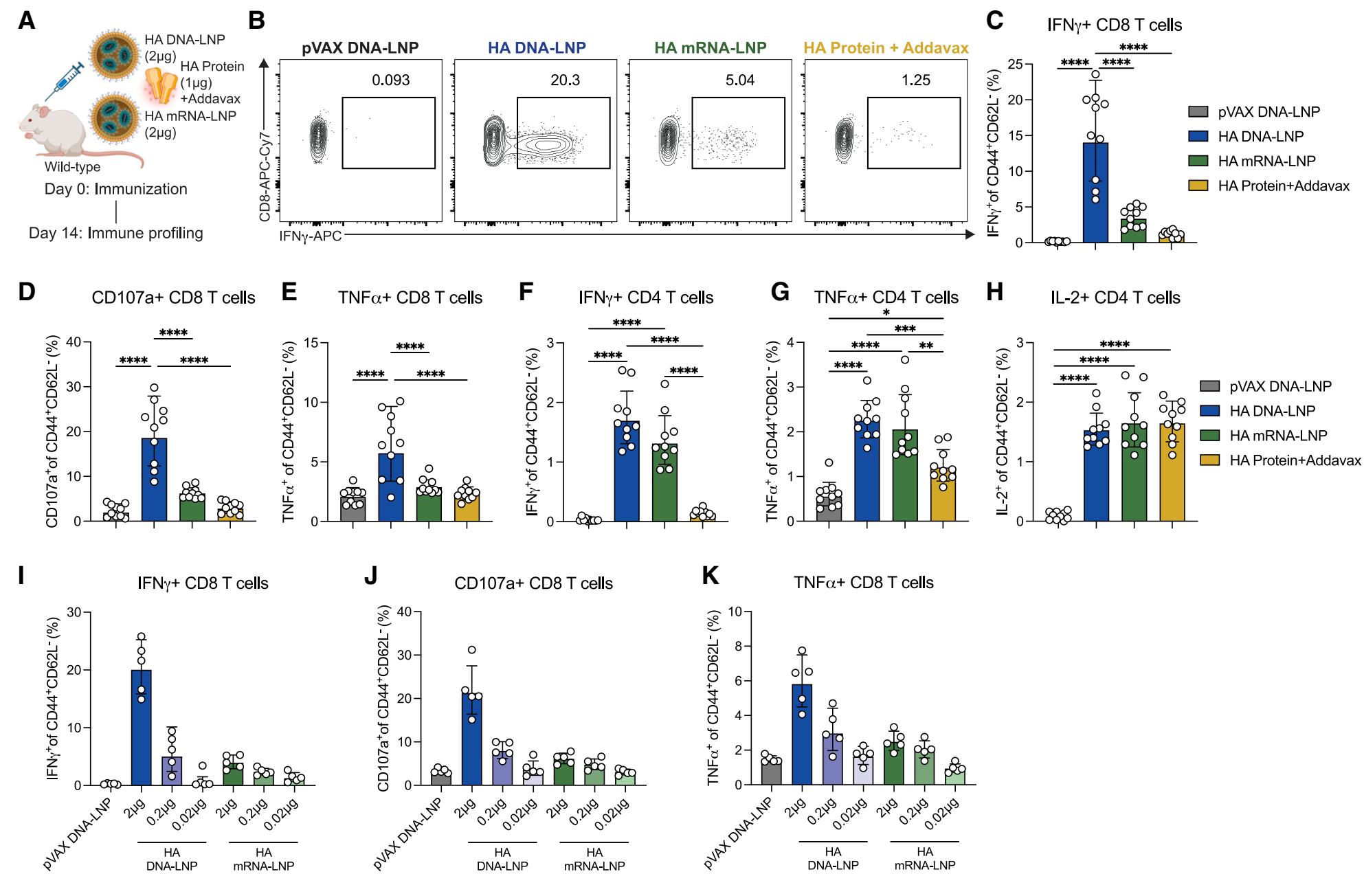
CD8+ T细胞反应:HA DNA-LNP诱导的IFNγ分泌CD8+ T细胞反应显著高于mRNA-LNP和佐剂蛋白组,分别高出3倍和7倍。在低剂量(0.2 mg)下,HA DNA-LNP诱导的CD8+ T细胞反应与高剂量(2 mg)的mRNA-LNP相当,显示出显著的剂量节省效应。 CD4+ T细胞反应:HA DNA-LNP和mRNA-LNP诱导的IFNγ分泌CD4+ T细胞反应相当,且显著高于佐剂蛋白组。然而,所有组诱导的IL-2分泌CD4+ T细胞反应相似。 |
|||||
|
Figure 3. HA DNA-LNP elicits potent antigen-specific CD8+ and CD4+ T cell responses |
|||||
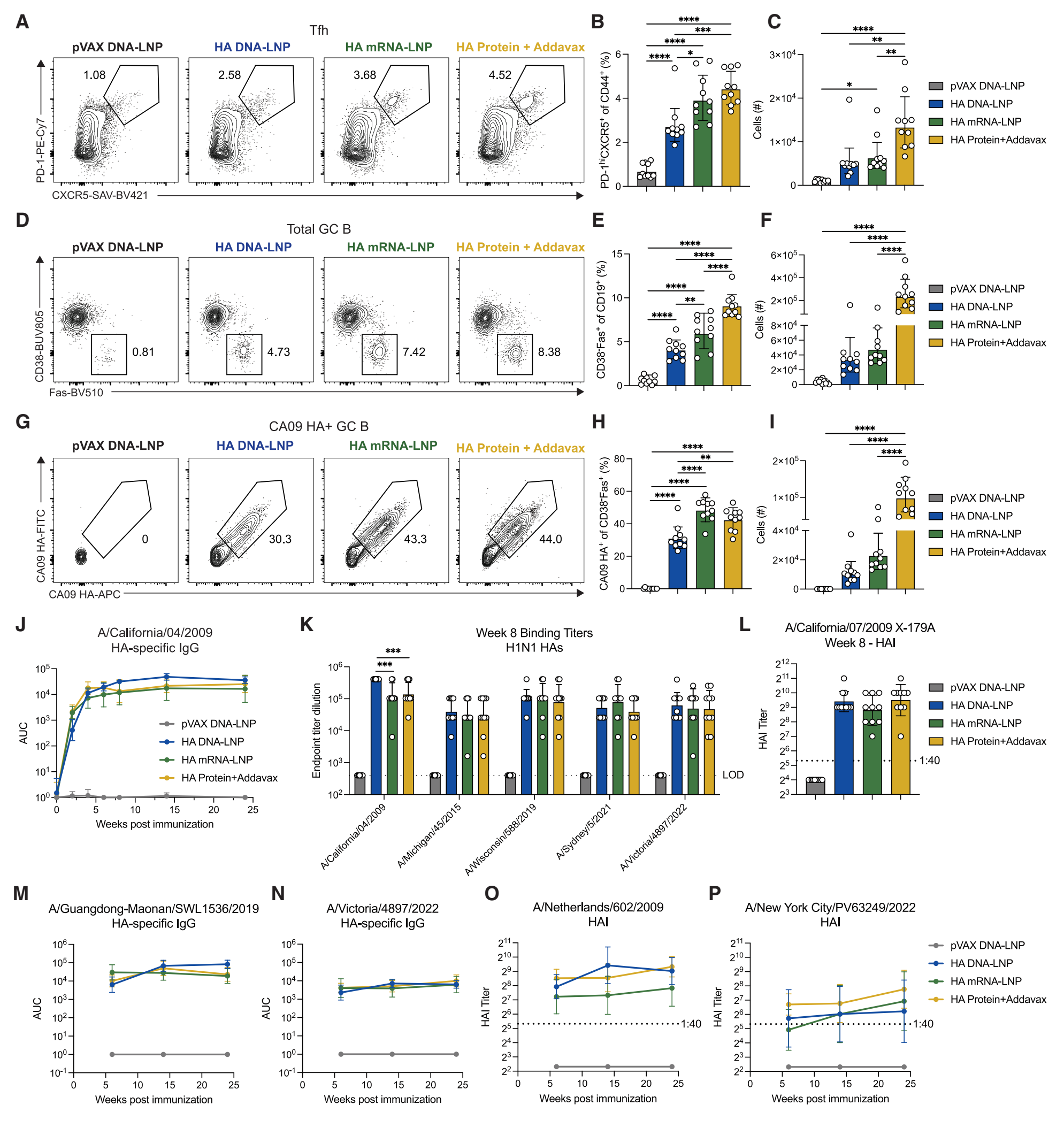
| 5. HA DNA-LNP诱导GC反应和功能性血清抗体: | |||||
|
GC反应:尽管HA DNA-LNP诱导的GC Tfh细胞频率有所下降,但GC B细胞的绝对数量与mRNA-LNP组相当。HA DNA-LNP诱导的抗原特异性GC B细胞反应在频率和绝对数量上均低于mRNA-LNP组。 血清抗体反应:HA DNA-LNP诱导的血清IgG反应与mRNA-LNP和佐剂蛋白组相当,且抗体滴度持久存在至少24周。对相关流感病毒株的抗体结合和血凝抑制(HAI)滴度也稳定存在24周。 |
|||||
|
Figure 4. HA DNA-LNP induces robust GC and serum responses |
|||||
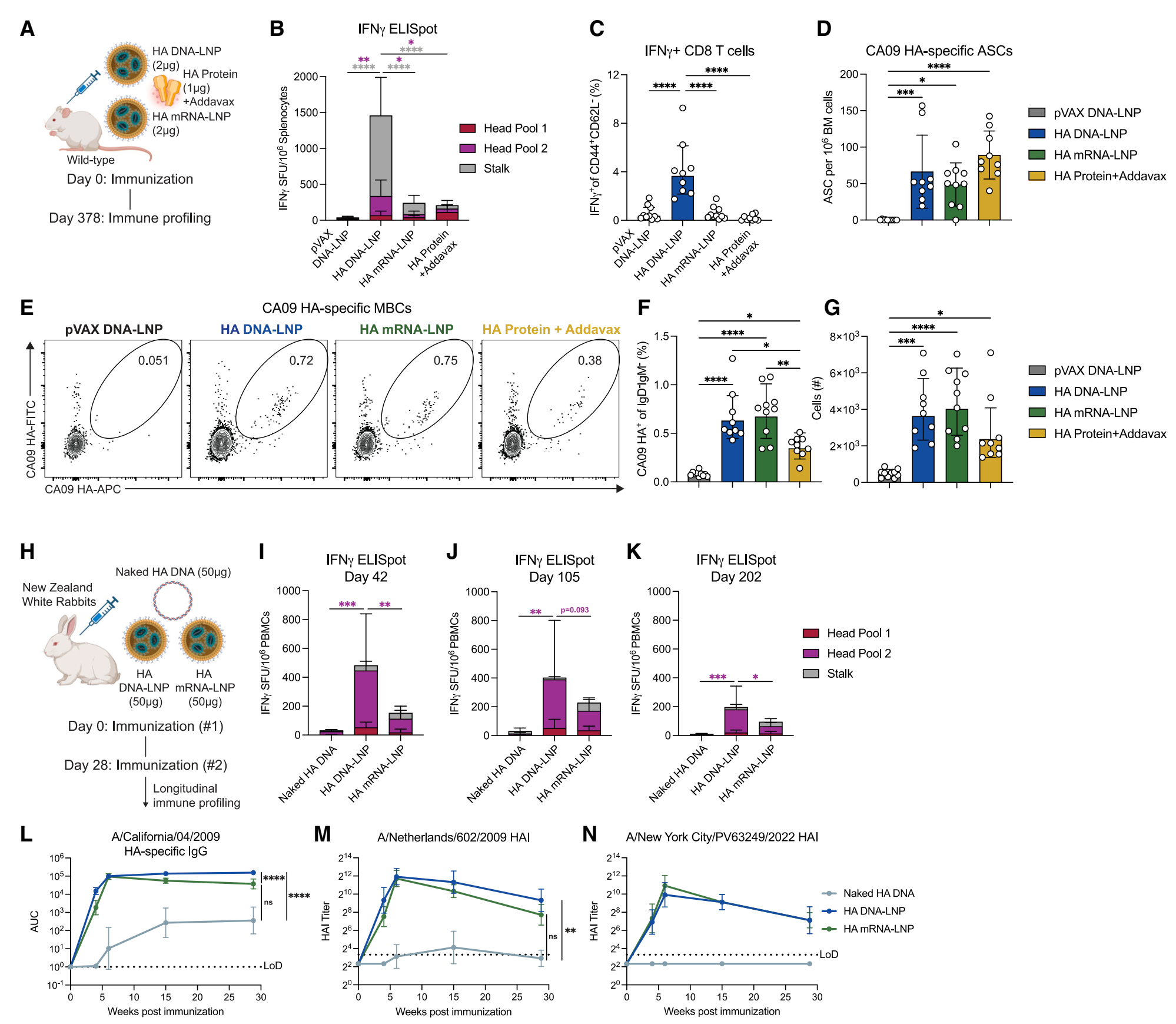
| 6. HA DNA-LNP在小鼠和兔子中诱导持久的记忆反应: | |||||
|
小鼠T细胞记忆反应:免疫后一年,HA DNA-LNP诱导的IFNγ分泌细胞反应比mRNA-LNP或佐剂蛋白组高约4倍。效应记忆CD8+ T细胞反应也显著高于其他组。 小鼠B细胞记忆反应:HA DNA-LNP诱导的骨髓驻留浆细胞反应与mRNA-LNP和佐剂蛋白组相当。抗原特异性记忆B细胞反应在HA DNA-LNP和mRNA-LNP组中相似,且显著高于佐剂蛋白组。 兔子T细胞反应:在兔子模型中,相同剂量下,HA DNA-LNP诱导的IFNγ分泌细胞反应在免疫后42天时比HA mRNA-LNP强约2倍,且在202天内仍能维持较高水平的反应。 兔子体液免疫反应:HA DNA-LNP和mRNA-LNP诱导的血清IgG反应相当,且对不同流感病毒株的HAI滴度也相似。尽管HA mRNA-LNP组的IgG反应有所减弱,但HA DNA-LNP组的反应保持稳定。 |
|||||
|
Figure 5. HA DNA-LNP induces potent memory responses in mice and rabbits |
|||||
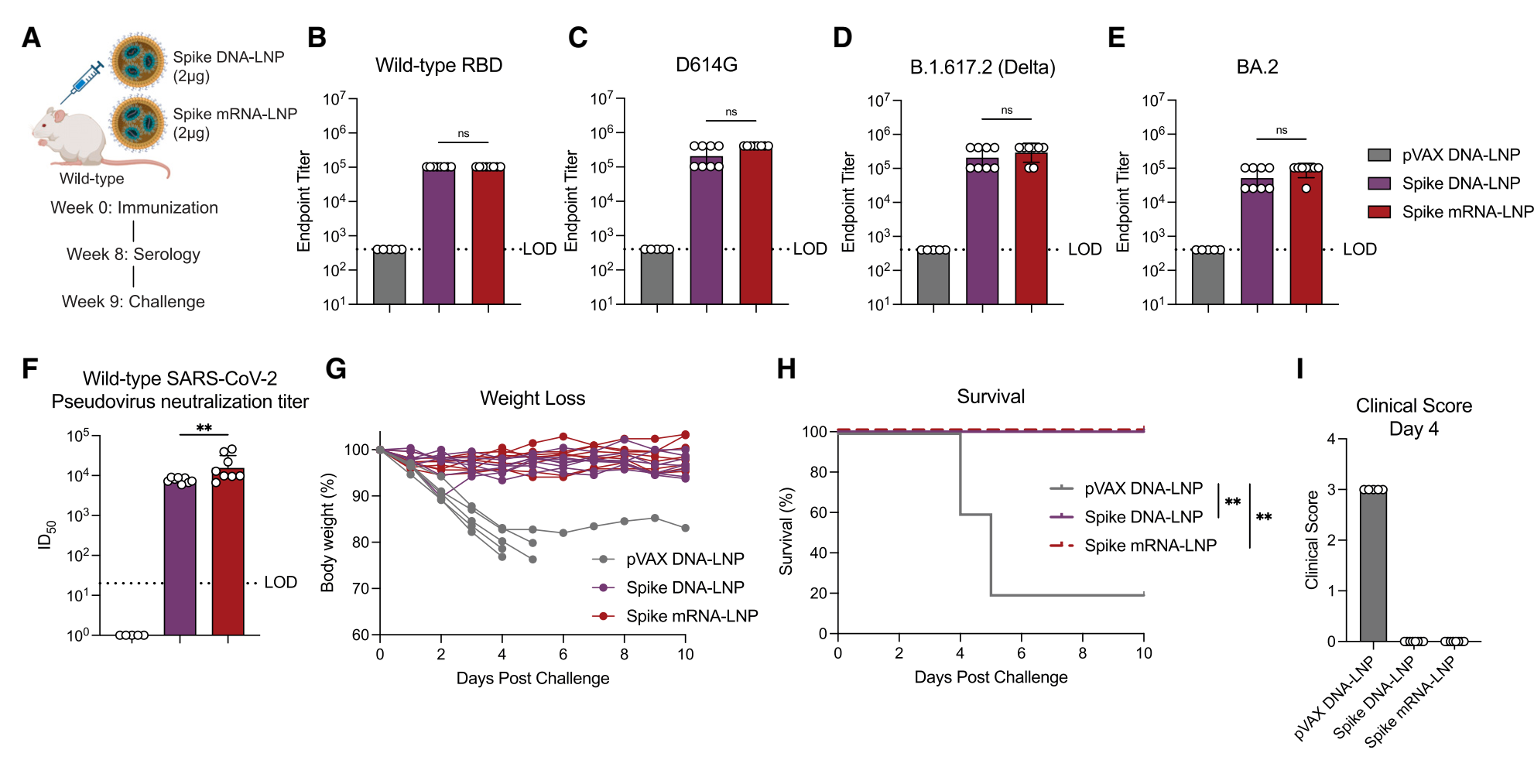
| 7. SARS-CoV-2刺突DNA-LNP在小鼠中的保护效力: | |||||
|
抗体反应:编码SARS-CoV-2刺突蛋白的DNA-LNP和mRNA-LNP疫苗在小鼠中诱导了相似的抗原特异性总IgG反应,对RBD和不同变异株的刺突蛋白均产生竞争性的抗体反应。 中和滴度:Spike DNA-LNP诱导的中和滴度与Spike mRNA-LNP相当。 保护效力:在小鼠攻毒实验中,Spike DNA-LNP和mRNA-LNP均能保护小鼠免受致死性SARS-CoV-2攻击,体重无明显下降且全部存活,而空载体对照组小鼠则出现体重下降和高死亡率。 |
|||||
|
Figure 6. SARS-CoV-2 spike DNA-LNP elicits serum antibody responses and is protective in lethal challenge |
|||||
结论 |
|||||
|
本研究描述了一种脂质纳米颗粒包裹的质粒DNA(DNA-LNP)制剂,该制剂能够诱导强大的先天免疫和适应性免疫反应,其血清学效力与mRNA-LNP和佐剂蛋白疫苗相当。通过使用编码流感血凝素(HA)的构建体,研究发现HA DNA-LNP免疫能够通过刺激干扰素基因刺激因子(STING)依赖的方式上调并激活迁移性树突状细胞(DC)亚群。与mRNA-LNP或佐剂蛋白相比,HA DNA-LNP诱导了更优越的抗原特异性CD8+ T细胞反应,且记忆反应可持续超过一年。在兔子模型中,相同剂量下,HA DNA-LNP的免疫反应与mRNA-LNP相当或更优。此外,编码SARS-CoV-2刺突蛋白的DNA-LNP在小鼠模型中显示出与刺突mRNA-LNP相当的保护效力。本研究揭示了DNA-LNP平台特有的启动机制,为预防性和治疗性疫苗的开发提供了新途径。 |
|||||
| 参考文献:Tursi, Nicholas J., et al. "Modulation of lipid nanoparticle-formulated plasmid DNA drives innate immune activation promoting adaptive immunity." Cell Reports Medicine 6.4 (2025). | |||||