产品









服务
联系我们
400-021-7882
021-54281913
021-54281913
|
结直肠癌是全球癌症死亡的第二大原因,早期结直肠癌可通过手术治疗,但复发和耐药性问题亟待解决。癌症免疫治疗通过激活患者自身免疫系统对抗恶性细胞,展现出革命性潜力。PCVs通过靶向患者特异性新抗原,诱导特异性抗肿瘤免疫反应,成为研究热点。然而,新抗原的准确预测和疫苗设计的优化仍是主要挑战。近日,韩国加图立大学Jae-Hwan Nam团队在Advanced Science上发表名为“Rapid-Turnaround Co-Administration of mRNA-Based MHC-I and MHC-II-Restricted Neoantigens Enhances Immune Responses of Antigen-Specific CD8+ T Cells and Anti-Cancer Efficacy in Colorectal Cancer”的研究,阐述了抗原序列选择的重要性。 |
|||||
|
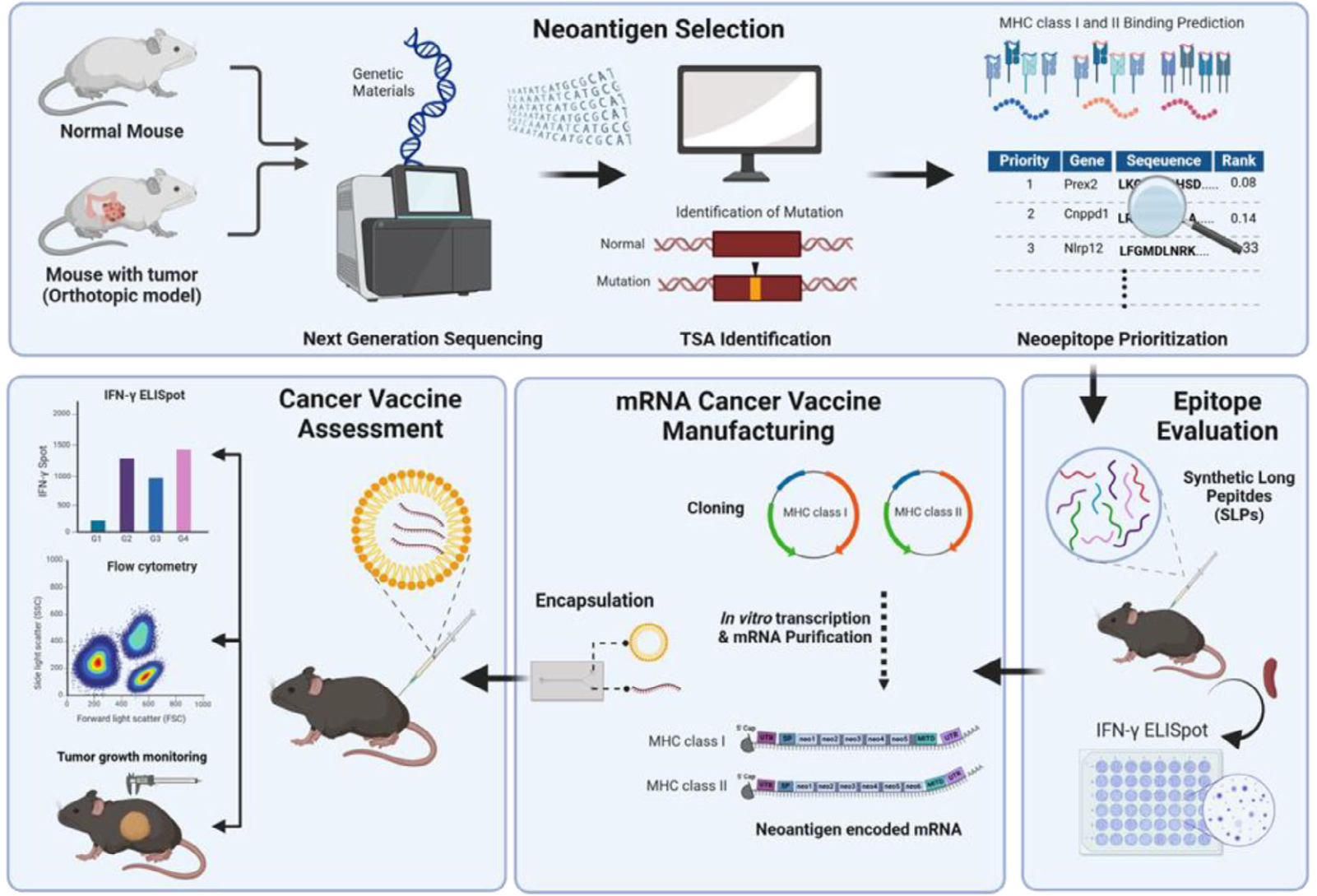
Figure 1. Schematic illustration of neoantigen validation |
|||||
01
|
|||||
| 1.1 新抗原预测: | |||||
|
在常规个性化癌症疫苗(PCV)新抗原设计中,通常通过靶向癌症组织中特有的体细胞突变来设计表位。本研究利用MC38细胞系构建了一个原位模型,利用该细胞系与小鼠品系之间的基因组差异来诱导肿瘤特异性免疫反应(图1a)。通过两种变异检测工具(variant callers)识别出仅存在于肿瘤中的变异,最终筛选出546个单核苷酸变异(SNVs)和68个插入/缺失(InDels)作为新抗原设计的候选序列。 此外,通过比较正常和肿瘤样本的mRNA-seq数据,使用ARRIBA和STAR fusion工具分析了基因融合,最终确定了6个基因融合作为新抗原设计的候选序列。经过综合考虑MHC结合能力、突变表达水平以及与自身抗原的独特性,最终设计了20个MHC-I和12个MHC-II限制性新抗原。 |
|||||
| 1.2 预测新抗原的免疫原性: | |||||
|
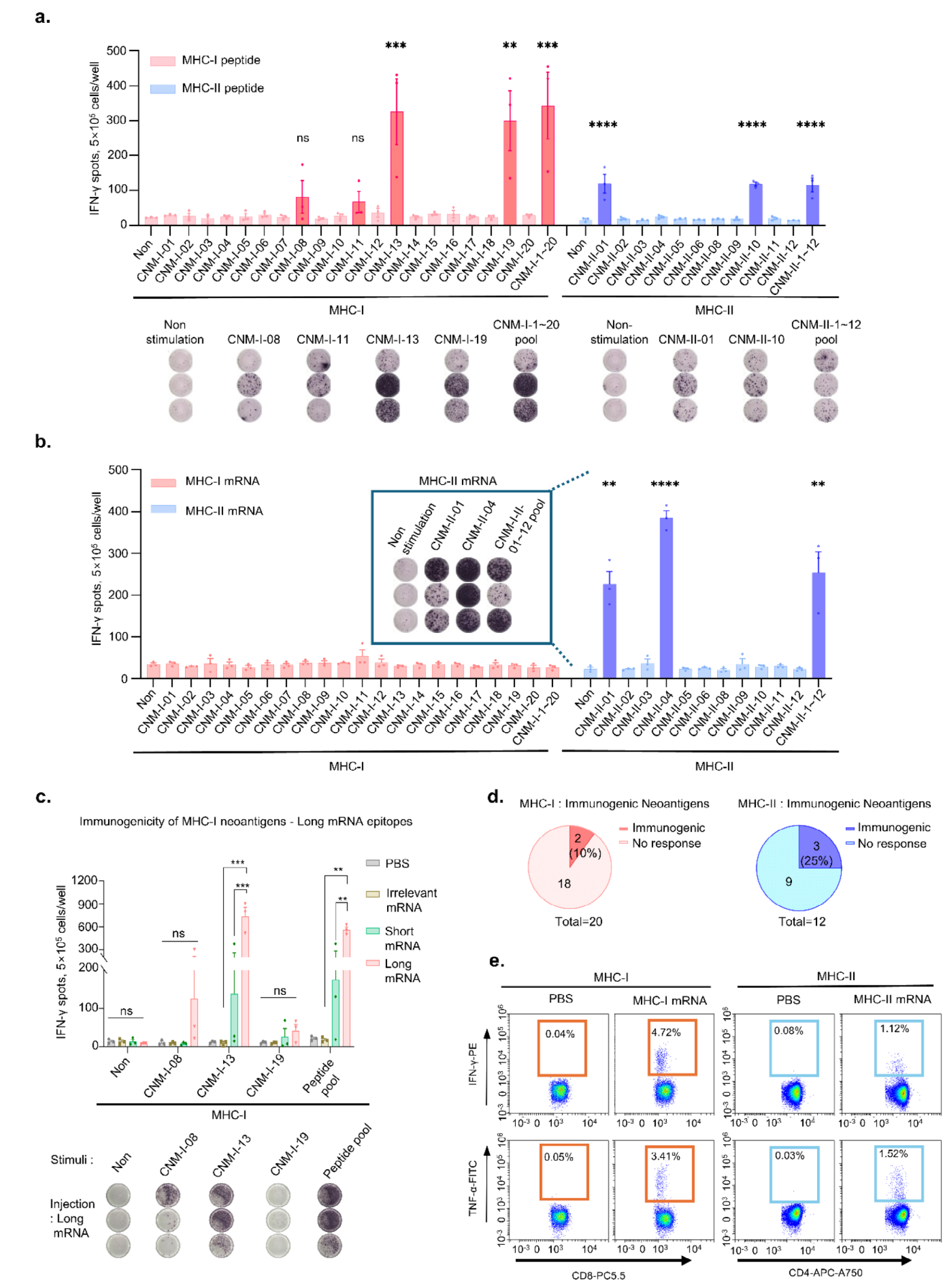
为了评估预测新抗原的免疫原性,将32个具有高MHC-I和MHC-II结合预测评分的新抗原候选序列合成为合成长肽(SLPs)。其中,20个与MHC-I相关的新抗原被合成为8-10个氨基酸的肽,而12个与MHC-II相关的新抗原被合成为27个氨基酸的肽。这些肽与poly I:C混合后,每周一次皮下注射给C57BL/6小鼠。最后一次免疫后一周,收集脾细胞进行IFN-γ ELISpot分析。结果显示,两个MHC-I相关的新抗原(CNM-I-13和CNM-I-19)和两个MHC-II相关的新抗原(CNM-II-01和CNM-II-10)表现出显著的抗原特异性IFN-γ分泌(图2a)。 随后,构建了编码相应新抗原的mRNA,并封装在脂质纳米颗粒(LNPs)中。将mRNA疫苗在0、4、7和14天皮下注射给C57BL/6小鼠。编码MHC-II限制性新抗原的mRNA疫苗(CNM-II-01和CNM-II-04)显著增加了IFN-γ分泌,而编码MHC-I相关新抗原的mRNA疫苗在治疗期间未诱导IFN-γ分泌(图2b)。进一步构建了编码长序列(27个氨基酸)MHC-I相关新抗原的mRNA疫苗,发现其比短序列(8-10个氨基酸)mRNA疫苗更具免疫原性(图2c)。总体而言,通过计算机模拟方法预测的新抗原中,10%的MHC-I抗原和25%的MHC-II抗原表现出免疫原性(图2d)。 流式细胞术分析证实,预测的MHC-I和MHC-II抗原途径分别诱导了细胞因子分泌。MHC-I新抗原诱导了CD8+细胞特异性的IFN-γ和肿瘤坏死因子-α(TNF-α)分泌,而MHC-II新抗原则促使CD4+细胞特异性的IFN-γ和TNF-α分泌(图2e)。 |
|||||
|
Figure 2. Immunogenicity identification of predicted neoantigen candidates. |
|||||
| 1.3 联合递送MHC-I和MHC-II抗原的mRNA对新抗原特异性CD8+T细胞介导的免疫反应的影响: | |||||
|
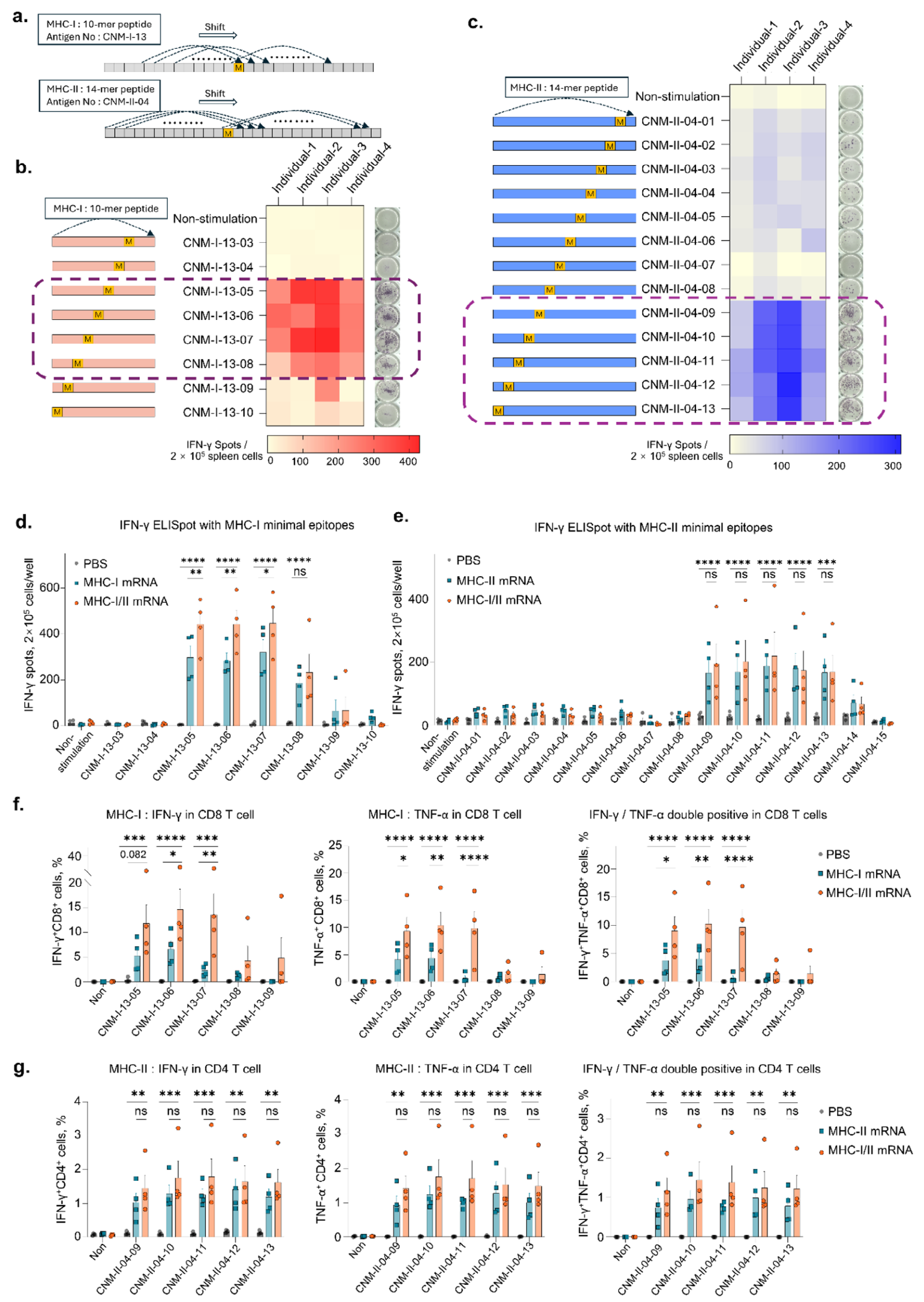
为了确定最小表位,设计了MHC-I抗原(CNM-I-13)内的10-mer肽(CNM-I-13-01至CNM-I-13-10)和MHC-II抗原(CNM-II-04)内的14-mer肽(CNM-II-04-01至CNM-II-04-13),包括每个突变(图3a)。发现CNM-I-13-05、-06、-07和-08在MHC-I限制性抗原中具有高度免疫原性(图3b),而CNM-II-04-09、-10、-11、-12和-13在MHC-II限制性抗原中强烈诱导抗原特异性IFN-γ分泌(图3c)。 通过流式细胞术分析确认,联合递送MHC-I和MHC-II疫苗显著增加了CD8+T细胞中IFN-γ或TNF-α的分泌,但在CD4+T细胞中未观察到显著差异(图3f,g)。这些结果表明,联合递送MHC-I和MHC-II不仅触发了各自的抗原特异性免疫反应,而且MHC-II抗原刺激的CD4+T细胞活化有助于抗原特异性CD8+T细胞的活化。 |
|||||
|
Figure 3. Defining the minimal epitopes for MHC-I and MHC-II neoantigens. |
|||||
| 1.4 选定的MHC-I/II抗原基础mRNA疫苗的抗癌效果: | |||||
|
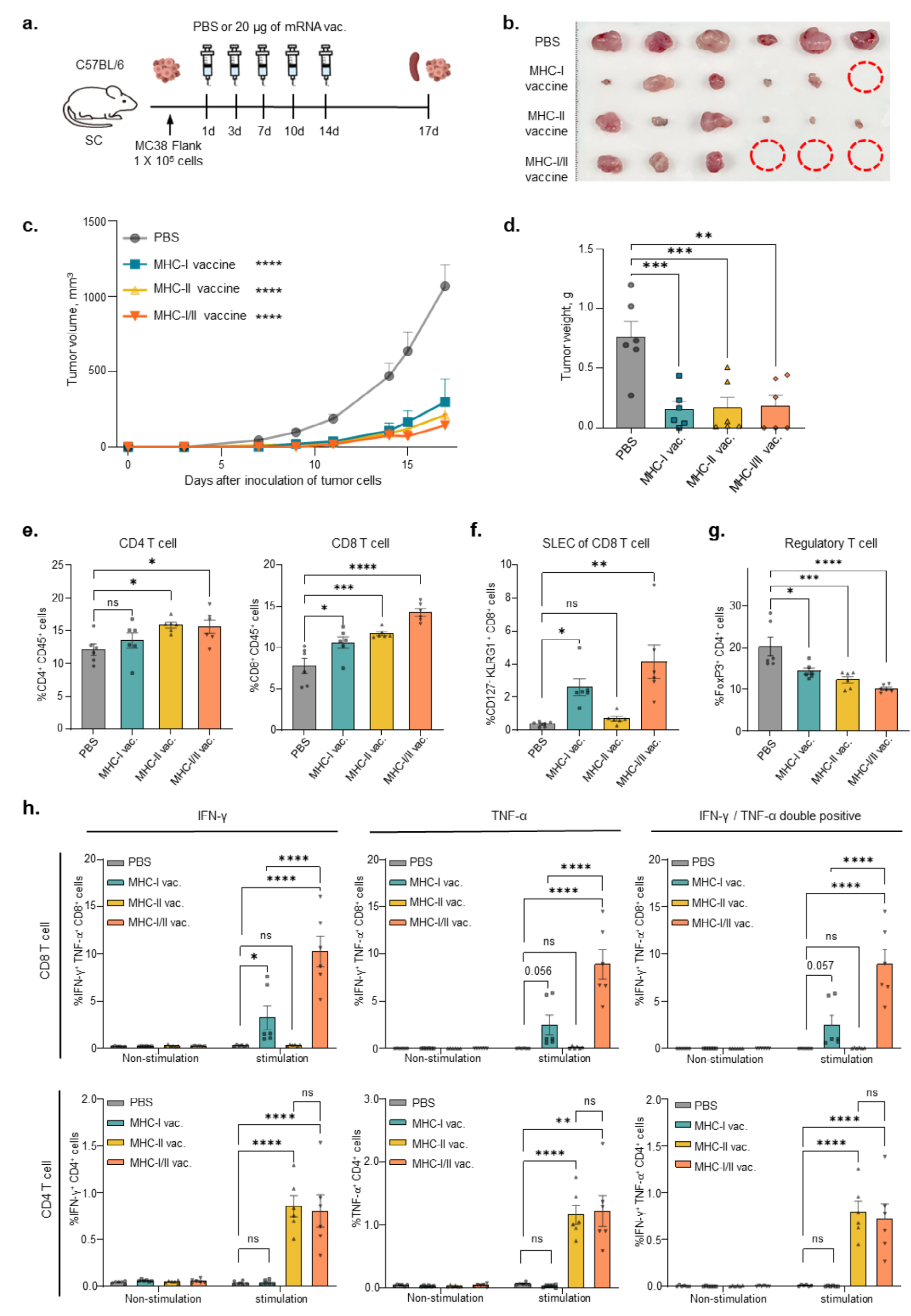
在MC38肿瘤小鼠模型中,分别给予MHC-I疫苗、MHC-II疫苗或两者联合疫苗,评估mRNA疫苗的抗癌效果(图4a)。与磷酸盐缓冲液(PBS)处理组相比,mRNA疫苗处理组显著抑制了结肠肿瘤的生长和体积(图4b-d)。联合递送MHC-I和MHC-II疫苗组无瘤小鼠数量显著增加(3/6),高于单独给予MHC-I(1/6)或MHC-II疫苗(0/6)组。此外,流式细胞术数据显示,MHC-II疫苗处理组CD4+/CD8+T细胞数量显著增加(图4e)。联合递送MHC-I和MHC-II疫苗比单独给予MHC-I疫苗诱导了更多的CD8+短寿命效应T细胞(SLECs,CD127-KLRG1+的CD8+细胞)增殖(图4f)。所有疫苗处理组的调节性T细胞(Treg)增殖均显著减少(图4g)。 在肿瘤小鼠模型中,联合递送MHC-I和MHC-II疫苗组的抗原特异性CD8+T细胞分泌的IFN-γ和TNF-α水平显著高于单独给予MHC-I疫苗组,而CD4+T细胞的IFN-γ和TNF-α分泌水平在MHC-II疫苗处理组和联合递送组之间无显著差异(图4h)。 |
|||||
|
Figure 4. Co-administration with MHC-I and MHC-II-restricted neoantigens enhanced anti-cancer efficacy via cell-mediated immune responses |
|||||
| 1.5 mRNA疫苗减少术后肿瘤复发并在小鼠结肠癌模型中预防性接种: | |||||
|
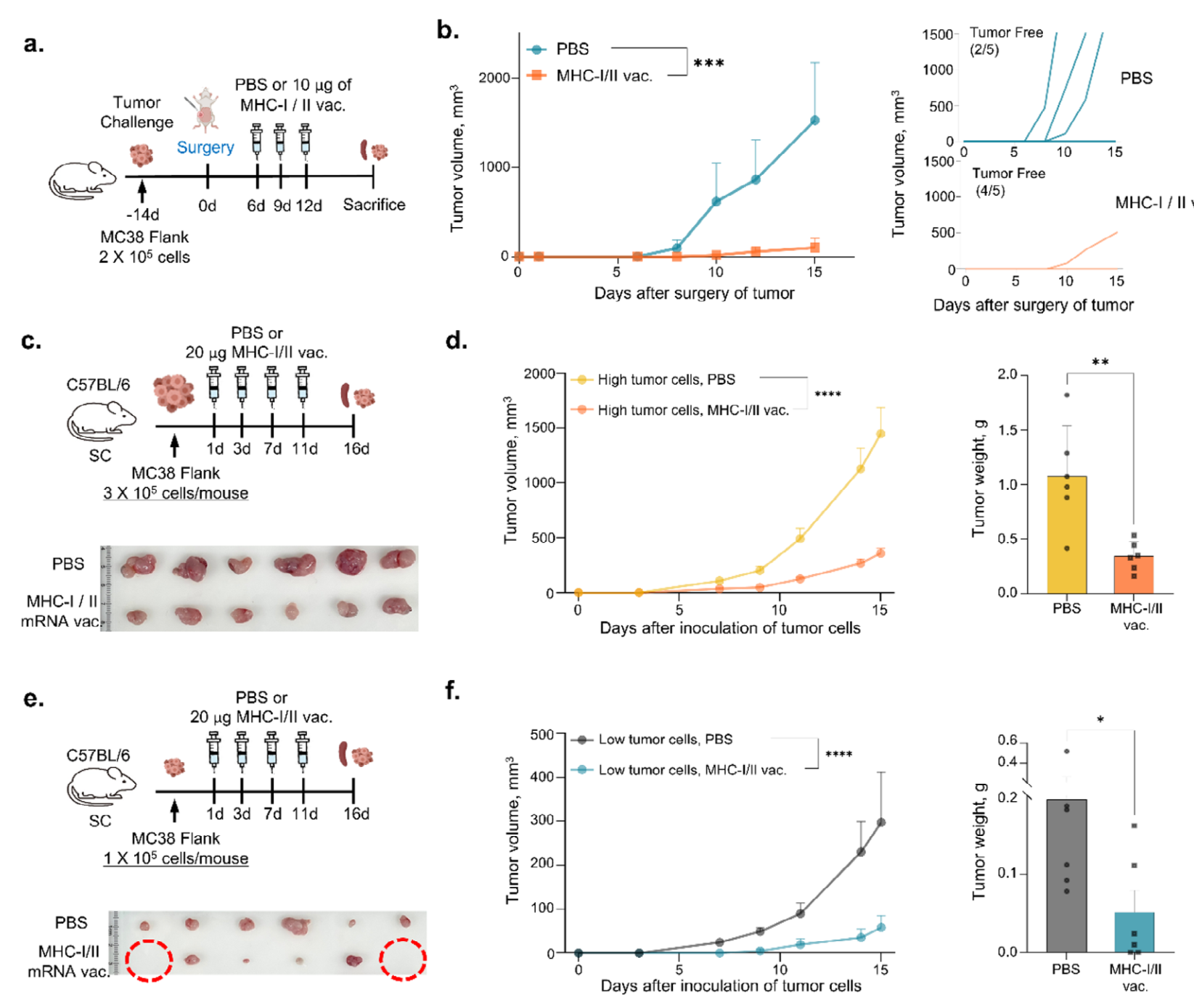
为了测试mRNA疫苗在预防肿瘤复发方面的效果,对肿瘤小鼠进行手术切除,并注射PBS或联合递送MHC-I和MHC-II mRNA疫苗(图5a)。与PBS处理组相比,mRNA疫苗处理组肿瘤复发显著减少(图5b)。此外,通过每周一次给予PBS或MHC-I/MHC-II疫苗,持续三周,测试了预防性接种的抗癌效果。与对照组相比,接受mRNA基础新抗原疫苗处理的小鼠肿瘤生长显著减少,无瘤小鼠数量增加。IFN-γ ELISpot分析确认了预防性接种后新抗原特异性IFN-γ分泌,表明产生了强烈的免疫反应。 |
|||||
| 1.6 mRNA疫苗在结肠癌早期发展中的抗癌效果: | |||||
|
通过接种不同数量的MC38细胞(1×105至3×105个细胞)给C57BL/6小鼠,评估mRNA疫苗在不同癌症发展阶段的抗癌效果(图5c-f)。与PBS处理组相比,疫苗处理组肿瘤体积和重量显著减少,且在接种低剂量癌细胞的小鼠中抗肿瘤效果更显著(图5c-f)。特别是,mRNA疫苗处理组中有两只小鼠无瘤(图5e)。细胞毒性测定显示,从mRNA疫苗处理小鼠中分离的肿瘤浸润淋巴细胞(TILs)对MC38-luc结肠癌细胞的杀伤作用显著增强。流式细胞术和免疫组化分析显示,mRNA疫苗处理组肿瘤组织中CD8+T细胞数量显著增加,且这些细胞浸润肿瘤并影响肿瘤细胞死亡。 |
|||||
|
Figure 5. mRNA vaccine reduces tumor recurrence after surgery in a mouse colon cancer model and increase anti-cancer efficacy in early colon cancer development. |
|||||
| 1.7 基于mRNA疫苗治疗时间的抗癌效果: | |||||
|
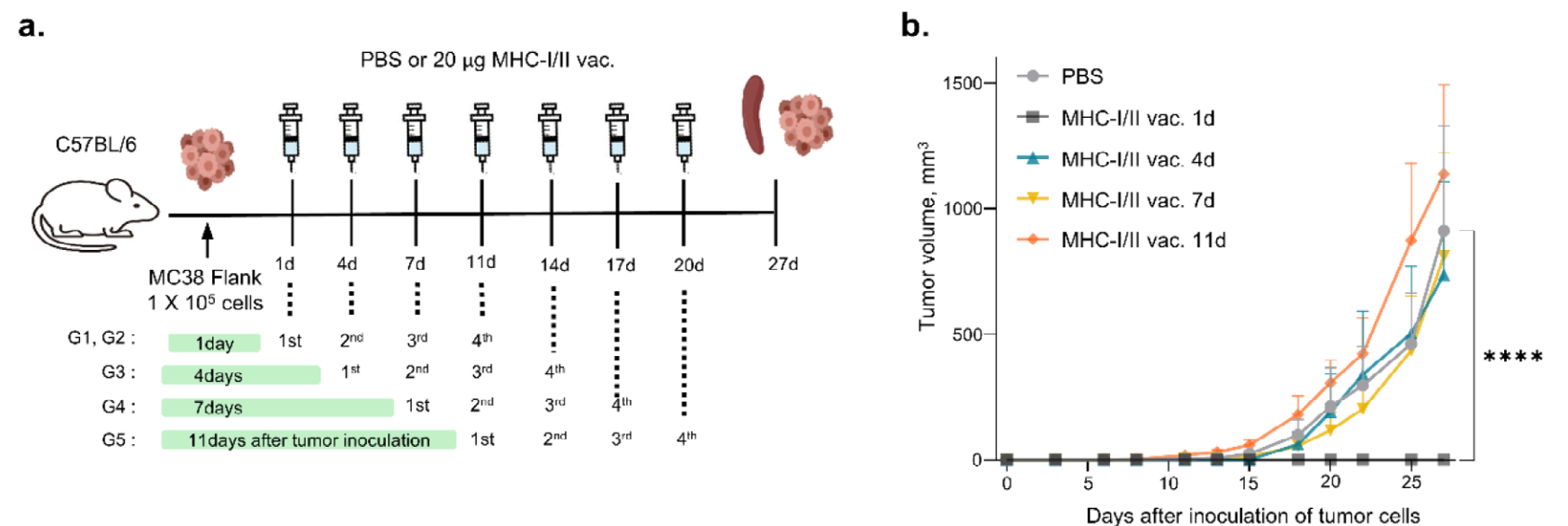
在MC38肿瘤小鼠模型中,分别在接种肿瘤细胞后的第1、4、7和11天给予主要mRNA疫苗接种,以确认基于接种时间的癌症疫苗效果(图6a)。与对照组相比,最早接种mRNA疫苗的小鼠肿瘤生长显著抑制,且七只小鼠中有六只肿瘤消失(图6b)。通过皮尔逊相关系数评估抗原特异性T细胞相关IFN-γ分泌与肿瘤大小之间的相关性,结果显示负相关(r=-0.835,p<0.001)。流式细胞术分析T细胞耗竭标记物显示,早期免疫组小鼠CD4+或CD8+T细胞中PD-1和Tim-3表达较低。随着接种时间的延迟,CD4+或CD8+T细胞中Tim-3表达逐渐增加。这些结果表明,早期接种mRNA癌症疫苗并与免疫检查点抑制剂(ICIs)联合治疗对于有效的抗癌治疗是必要的。 |
|||||
|
Figure 6. Early-stage vaccination produces a robust anti-cancer effect. |
|||||
| 1.8 mRNA疫苗接种诱导持续的MHC-I和MHC-II抗原特异性免疫反应并减少肿瘤复发负担: | |||||
|
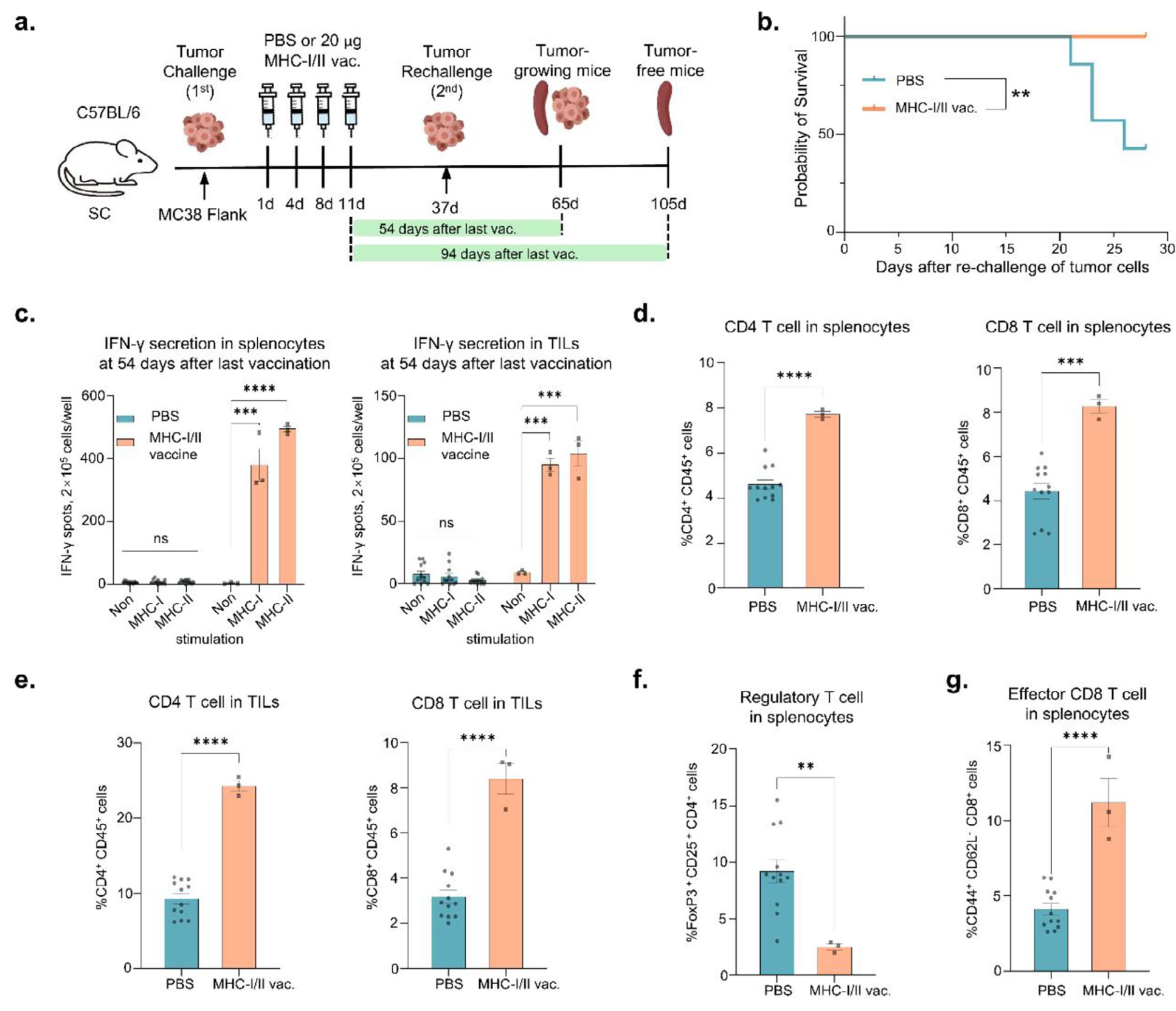
为了确定mRNA疫苗是否能通过诱导记忆T细胞分化来预防肿瘤复发或转移,对初次mRNA疫苗接种后无瘤的小鼠重新接种结肠癌细胞(图7a)。在mRNA疫苗处理组中,所有小鼠在重新接种后30天内均存活,而PBS处理的对照组小鼠存活率显著降低(图7b)。最后一次接种后54天,收集脾细胞和TILs进行IFN-γ ELISpot分析,确认mRNA疫苗诱导的抗原特异性免疫反应的持久性(图7c)。此外,mRNA疫苗处理增加了脾脏和TILs中CD4+或CD8+T细胞的数量(图7d和7e)。在最后一次接种后54天,mRNA疫苗处理的小鼠脾脏中Treg细胞(FoxP3+CD25+CD4+细胞)减少,同时效应记忆CD8+T细胞(CD44+CD62L-CD8+细胞)增加(图7f,g),表明mRNA疫苗通过产生记忆T细胞维持长期免疫反应。此外,最后一次接种后94天,脾细胞中的新抗原特异性T细胞反应仍然存在(图7h)。因此,mRNA疫苗接种在脾细胞和TILs中诱导了持续的MHC-I和MHC-II抗原特异性免疫反应。 |
|||||
|
Figure 7. mRNA vaccines prevent tumor recurrence and induced persistent MHC-I and MHC-II antigen-specific immune responses. |
|||||
| 1.9 mRNA疫苗与免疫检查点抑制剂联合治疗的抗癌协同效应: | |||||
|
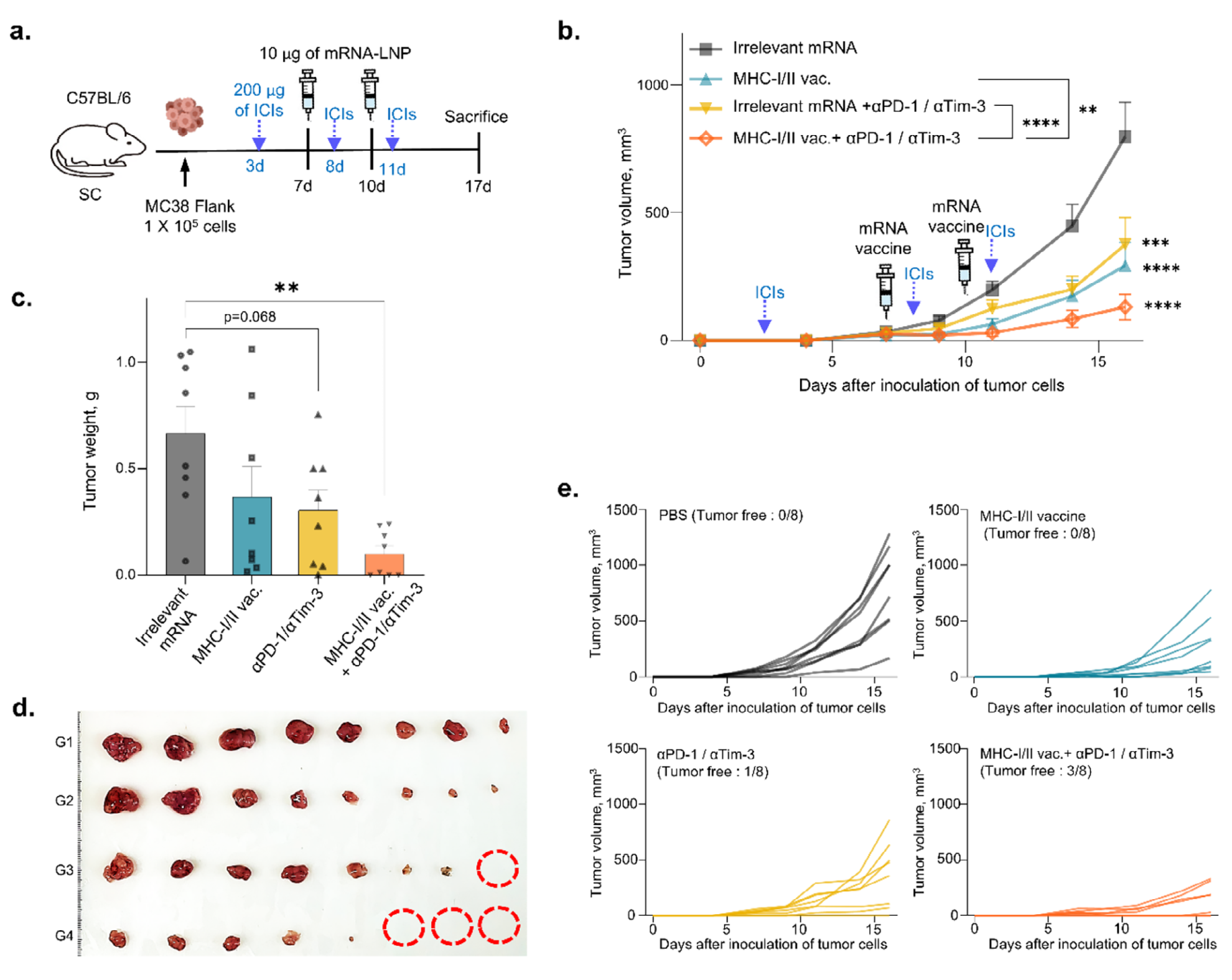
鉴于mRNA疫苗增加了免疫检查点分子的表达,我们在结肠癌模型中进行了mRNA疫苗与ICIs的联合治疗。C57BL/6小鼠右侧接种1×10^5个MC38结肠癌细胞后,分别在第3、8和11天给予200μg ICIs(抗小鼠PD-1和Tim-3抗体),在第7和10天给予10μg mRNA疫苗,或两者联合(图8a)。值得注意的是,与对照组相比,mRNA疫苗和ICIs处理组的肿瘤生长显著减少(图8b,c)。联合治疗组的肿瘤生长抑制比单独使用mRNA疫苗或ICIs更为显著(图8b,c)。此外,在联合治疗组中,三只小鼠的肿瘤完全消退,突显了对肿瘤生长抑制的协同效应(图8d,e)。 |
|||||
|
Figure 8. Combination therapy of mRNA vaccines with immune checkpoint inhibitors synergistically inhibits tumor growth. |
|||||
02
|
|||||
|
本研究开发的mRNA基础PCV平台通过联合递送MHC-I和MHC-II限制性新抗原,展示了显著的抗癌效果。该平台不仅减少了术后复发,还通过与ICIs的联合治疗,显著抑制了肿瘤生长。本研究通过联合递送MHC-I和MHC-II限制性新抗原,显著增强了mRNA疫苗的抗癌效果。这一策略不仅激活了CD8+T细胞的直接杀伤作用,还通过CD4+T细胞的辅助作用,优化了抗肿瘤免疫应答。早期疫苗接种和与ICIs的联合治疗进一步提高了疗效,为临床应用提供了重要参考。 |
|||||
| 参考文献:Cho, Seongje, et al. "Rapid‐Turnaround Co‐Administration of mRNA‐Based MHC‐I and MHC‐II‐Restricted Neoantigens Enhances Immune Responses of Antigen‐Specific CD8+ T Cells and Anti‐Cancer Efficacy in Colorectal Cancer." Advanced Science (2025): e06426. | |||||